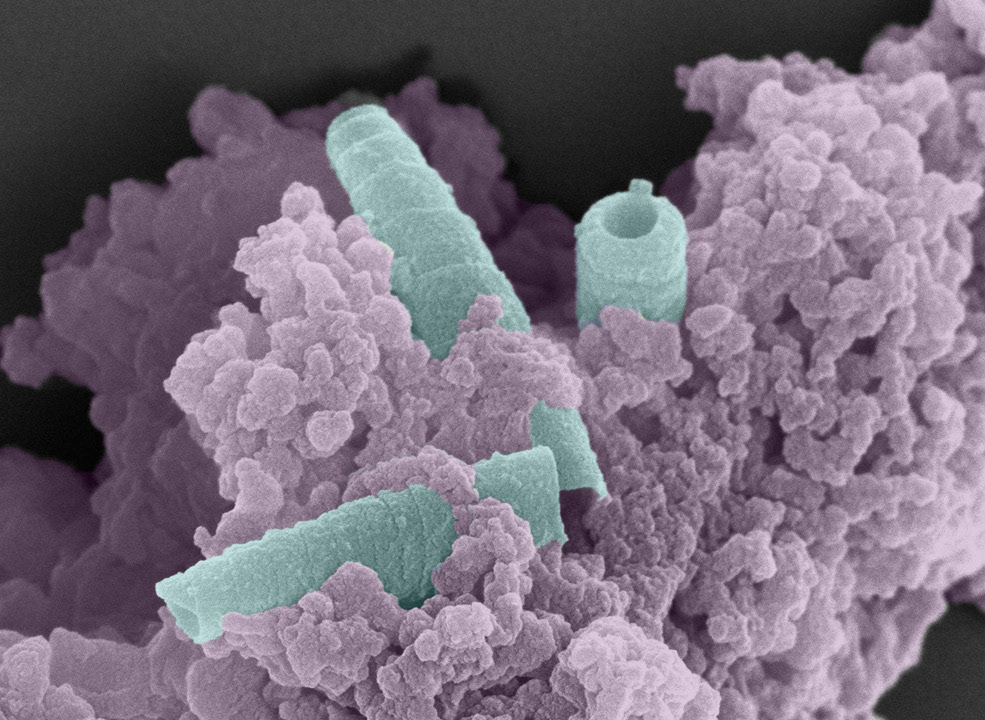
Production of the Pseudomonas aeruginosa R-body virulence factor and its roles in interkingdom interactions
P. aeruginosa is an opportunistic pathogen that colonizes diverse hosts and is a major cause of recalcitrant infections in humans. P. aeruginosa isolates show broad variation in their degree of toxicity in infection models, and strain surveys and genomic analyses have identified sequences that correlate with high pathogenicity. We have found that one of these “virulence-conferring” regions codes for the production of R-bodies, coiled intracellular protein ribbons that can extend to form large javelin-like structures.
Until recently, detailed studies of R-body production had been carried out for only two other types of bacteria—a parasite of unicellular eukaryotes and a plant symbiont—which are not closely related to P. aeruginosa. Yet phylogenetic analyses indicate that diverse proteobacteria, including several species of pseudomonads, have the capacity to produce R-bodies. In aquatic and soil settings, R-bodies may contribute to interkingdom interactions, for example by killing unicellular eukaryotes that prey on bacteria. We have shown that P. aeruginosa PA14 R-bodies contribute to plant host colonization and to pathogenicity in nematode worms, suggesting that these structures may have been co-opted to play roles in opportunistic infections.
Our foundational characterization of R-body production and toxicity in P. aeruginosa PA14 has provided exciting insights into the regulation of R-body structural genes, R-body assembly in vivo, and the mechanism of R-body toxicity during infection. The genes for R-body production are stochastically expressed, which makes sense for a product that extends to 5x the length of the bacterial cell. The locus conferring R-body production in PA14 contains homologs to R-body structural genes from unrelated bacteria, but also contains genes for R-body-associated proteins that are unique to P. aeruginosa. We hypothesize that these genes contribute to R-body assembly or to toxicity in the host.
Our working model of toxicity involves endocytosis of PA14 cells, or R-bodies themselves, by host cells. We hypothesize that R-body extension and piercing of the lysosomal membrane and/or concomitant release of a bacterial toxin triggers host translational inhibition and killing.
Outstanding questions include:
What are the molecular mechanisms controlling expression of R-body structural genes and R-body assembly?
What mechanisms limit R-body gene expression to <1% of the population?
What are the components of the R-body, how are they arranged in the coiled and extended forms?
Is pseudomonad R-body production toxic to predatory unicellular eukaryotes?
How does R-body production contribute to toxicity in the host, i.e., what is the mechanism of host damage?